Гадрозавриды[1][2], или гадрозавры[2] (лат. Hadrosauridae) — семейство так называемых «утконосых» динозавров из подотряда орнитопод, время жизни которых — меловой период.
| † Гадрозавриды | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Копия скелета Edmontosaurus annectens (Музей естественной истории Оксфордского университета) | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Клада: Клада: Клада: Клада: Отряд: Клада: Подотряд: † Цераподы Инфраотряд: Парвотряд: Надсемейство: Семейство: † Гадрозавриды |
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Hadrosauridae Cope, 1869 | |||||||||||||||||||||||||||||||||||||||||
| Геохронология Меловой период 145,0—66,0 млн лет
◄ Наше время
◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание
◄ Массовоепермское вымирание ◄ Девонское вымирание
◄ Ордовикско-силурийское вымирание
◄ Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
Самые ранние ископаемые остатки представителей семейства известны начиная с конца сантона и заканчиваются вместе с ископаемым материалом других динозавров в результате массового вымирания в конце маастрихта (мел-палеогеновое вымирание)[3]. Находки окаменелостей дают возможность говорить о широком географическое распространение данной группы динозавров, поскольку остатки гадрозавров обнаружены в Африке[4], Азии, Антарктиде, Европе, Северной и Южной Америке, за исключением Океании[3].
Характерной и отличительной особенностью семейства от других групп динозавров является строение носовой области, где кости были изменены в специализированные структуры, такие как черепные гребни у некоторых представителей[5]. Кроме того, они обладали сложной зубной системой, состоящей из нескольких зубных рядов, и ороговевшим клювом, что позволило им более эффективно питаться растительной пищей[6][7].
Большинство диагностических элементов скелета для данного семейства находятся в черепной области[8], на основе которых было описано большое количество видов. Тем не менее грудной и тазовый пояса также предоставляют широкий спектр таксономической информации[3]. По степени выраженности определённых морфологических особенностей гадрозавры делятся на два подсемейства — ламбеозаврины и гадрозаврины. В общей сложности на 2016 год было описано около 61 вида[9].
Описание
правитьДля семейства гадрозаврид характерен гребень различных форм. В этом гребне находились носовые полости, которые, вероятно, служили для издавания громких звуков. Дэвида Б. Вейсхампел и Коралия-Мария Жиану в книге «Трансильванские динозавры» предполагают, что эти наросты могли выполнять социальную функцию, но не могут утверждать, что это была форма полового диморфизма.

Гадрозавры имели плоский беззубый, схожий с утиным, кератиновый клюв, а в задней части челюсти находилось множество плоских зубов. Этот зубной аппарат, так называемые зубные «батареи», представляют собой комплекс из колонн постоянно сменяющихся на протяжении жизни зубов, способных перетирать самую жёсткую растительную пищу. Передвигались как на двух, так и на четырёх конечностях. Имели длинные задние и более короткие передние лапы. Длинный плоский хвост служил для равновесия[10]. Гадрозавриды вели стадный образ жизни, строили гнёзда, охраняли кладку и вместе заботились о потомстве.
Долгое время гадрозаврид, из-за строения передних конечностей с перепонками между 4-м и 5-м пальцами и плоским хвостом, считали полуводными водорослеядными животными. Сейчас же семейство «утконосых динозавров» относят к наземным обитателям. В желудке хорошо сохранившегося эдмонтозавра было найдено большое количество хвои, а не водорослей. Также установлено, что на передних конечностях у гадрозавров были копытца, а под хвостом имели мясистый нарост, из-за которого плавание было просто невозможным.
География находок
правитьАфрика
правитьДлительное время останки гадрозавров не находили на африканском континенте. В меловом периоде Африка была изолирована водой от европейского континента, и считалось что гадрозавры были неспособны достичь африканского континента. Однако в 2020 году, после описания нового рода Ajnabia, стало известно, что гадрозавры были распространены и на территории северной Африки. Предполагается, что миграция гадрозавров из подсемейства ламбеозаврин шла из Азии через Европу, и далее по морю в Африку[11]. Исследователи отмечают, что современные крупные наземные животные также могут преодолевать значительные расстояния по морю, активно плавая, будучи пассивно переносимы течениями или плывя на естественных плотах.
Северная Америка
правитьВ Мексике гадрозавры представляют собой самый обширный список находок среди всех групп динозавров, причем большинство сообщений поступает из северного региона. Несмотря на это, изучение мексиканских образцов было относительно скудным из-за фрагментарного состояния большинства образцов, в то же время число исследований гадрозавров в Мексике в последнее время увеличилось[3].
Всеобъемлющий обзор гадрозавров Мексики показал, что возраст геологических подразделений с остатками костной, технологической, покровной, зубной и оолитовой природы этих организмов, относится к альбу—маастрихту, обнаруженых на территории штатов Нижняя Калифорния, Сонора, Чиуауа, Коауила, Пуэбла и Мичоакан. Собранные данные позволяют признать сравнительно низкое таксономическое разнообразие по отношению к остальной части Северной Америки. В настоящее время по костным остаткам описано пять видов — Kritosaurus navajovious, Tlatolophus galorum, Magnapaulia laticaudus, Velafrons coahuilensis и Latirhinus uitstlani. Также идентифицирован один ихновид группы. Другие скелетные останки гадрозавров в стране идентифицированы лишь на фрагментарном уровне[12][13].
В 2022 году на территории Провинциального парка динозавров в канадской провинции Альберта найдены две крупные окаменелости — ступня и часть хвоста, покрытые окаменелой кожей. Останки мумифицировались и прекрасно сохранились. Найденный гадрозавр погиб в промежутке между 77-75 миллионами лет назад, за десять миллионов лет до того, как динозавры вымерли[14].
Южная Америка
правитьКостные остатки гадрозавров на южноамериканском континенте в настоящее время ограничены аргентинскими провинциями Ла-Пампа, Рио-Негро, Чубут и Санта-Крус, а также югом Чили[15].
О первых окаменелостях патагонских гадрозавров сообщил Р. Казамикела в 1964 году, который предположил, что их присутствие на юге континента было следствием события расселения из Северной Америки в конце мелового периода. Впоследствии эта гипотеза была подкреплена открытием более информативных и полных образцов[16]. С тех пор сведения о гадрозаврах в Южной Америке, и особенно в Патагонии, значительно расширились. Были описаны шесть различных таксонов: Secernosaurus koerneri, Kritosaurus australis, Bonapartesaurus rionegrensis, Lapampasaurus cholinoi, Willinakaqe salitralensis и Huallasaurus australis. Кроме того Родольфо Кориа сообщил о присутствии нескольких неописанных морфотипов гадрозавров из формации Аллен[17]. Наличие трех различных таксонов гадрозаврид из формации Аллен, плюс два неописанных морфотипа, привело к большим дебатам о таксономической обоснованности некоторых из этих динозавров, затрудняя отнесение видов к любому ранее известному таксону[15].
Филогенетическое положение патагонских гадрозавридов горячо обсуждается, и некоторые авторы поддерживают идею о том, что гадрозавры прибыли в Южную Америку по сухопутному мосту из Северной Америки, в то время как другие предполагают, что южноамериканские гадрозавры принадлежали к отдельной гондванской кладе[15].
Семейство гадрозаврид
править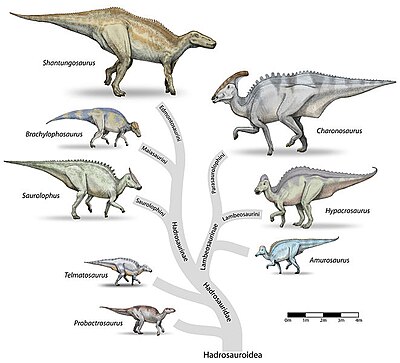
Семейство включает в себя два подсемейства:
- Ламбеозаврины (Lambeosaurinae) — подсемейство, характерной особенностью которых является полый гребень на голове[18].
- Гадрозаврины (Hadrosaurinae; другое название Saurolophinae) — подсемейство, представители которого обладали твёрдым гребнем на голове, или не имели гребня вовсе[18].
В некоторых кладистических анализах гадрозавров используется название Saurolophidae. Клада была впервые использована в 1928 году Францем Нопчей[19]. Данное название применяется редко, потому что клада включала в себя виды, которые формируют подсемейство завролофин (Saurolophinae) в семействе гадрозавров.
Тем не менее в 2010 году в результате филогенетического анализа Альберт Прието-Маркес обнаружил, что гадрозавр является сестринским таксоном для всех других гадрозаврид, и поэтому, соблюдая правила Международного кодекса зоологической номенклатуры, переименовал кладу гадрозаврин с твердым гребнем в Saurolophinae. Он вновь вернулся к употреблению клады Saurolophidae и определил её как группу, состоящую из последнего общего предка Saurolophus osborni и Lambeosaurus lambei, а также всех его потомков[20]. Хотя новое подсемейство Saurolophinae получило некоторое признание в научной литературе[21][22], не все современные палеонтологи восстанавливают гадрозавра отдельно от гадрозаврин[23][24]. В связи с этим название клады Hadrosaurinae практически является синонимом Saurolophinae, за исключением включения Hadrosaurus[25].
Классификация
править
По данным сайта Paleobiology Database, на декабрь 2020 года в семейство гадрозавров включают следующие вымершие роды и виды:[26]
- Роды incertae sedis
- Подсемейство Hadrosaurinae — Гадрозаврины
- Acristavus
- Anasazisaurus
- Anatotitan
- Augustynolophus
- Bonapartesaurus
- Brachylophosaurus
- Edmontosaurus
- Gryposaurus
- Hadrosaurus
- Kamuysaurus
- Kerberosaurus
- Kritosaurus
- Kundurosaurus
- Lophorhothon
- Maiasaura
- Mandschurosaurus
- Naashoibitosaurus
- Probrachylophosaurus
- Prosaurolophus
- Saurolophus
- Secernosaurus
- Shantungosaurus
- Ugrunaaluk
- Wulagasaurus
- Подсемейство Lambeosaurinae — Ламбеозаврины
- Adelolophus
- Adynomosaurus
- Ajnabia
- Amurosaurus
- Angulomastacator
- Aralosaurus
- Arenysaurus
- Blasisaurus
- Canardia
- Charonosaurus
- Corythosaurus
- Hadrosaurus paucidens
- Hypacrosaurus
- Jaxartosaurus
- Kazaklambia
- Koutalisaurus
- Lambeosaurus
- Magnapaulia
- Nipponosaurus
- Olorotitan
- Pararhabdodon
- Parasaurolophus
- Sahaliyania
- Tsintaosaurus
- Velafrons
Ниже показано филогенетическое древо, полученное в результате филогенетического анализа проведённого группой палеонтологов под руководством Себастьяна Розадиллы (2022), включая Huallasaurus и Kelumapusaura, которые были описаны в том же исследовании[15].
| Hadrosauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Примечания
править- ↑ Орнитоподы : [арх. 15 июня 2024] / Алифанов В. Р. // Океанариум — Оясио. — М. : Большая российская энциклопедия, 2014. — С. 463. — (Большая российская энциклопедия : [в 35 т.] / гл. ред. Ю. С. Осипов ; 2004—2017, т. 24). — ISBN 978-5-85270-361-3.
- ↑ 1 2 Алифанов В. Р., Кириллова И. В. Обзор дочетвертичных позвоночных острова Сахалин // Ученые записки Сахалинского государственного университета : сборник научных статей : [арх. 2 сентября 2017] / Гл. ред. и сост. Т. К. Злобин. — Южно-Сахалинск : СахГУ, 2008. — Вып. VII. — С. 40.
- ↑ 1 2 3 4 Reyna-Hernández, R.A., Rivera-Sylva, H.E., Silva-Martínez, L.E., and Guzman-Gutiérrez, J.R. 2021. A large hadrosaurid dinosaur from Presa San Antonio, Cerro del Pueblo Formation, Coahuila, Mexico. Acta Palaeontologica Polonica 66
- ↑ Longrich, Nicholas R.; Suberbiola, Xabier Pereda; Pyron, R. Alexander; Jalil, Nour-Eddine. The first duckbill dinosaur (Hadrosauridae: Lambeosaurinae) from Africa and the role of oceanic dispersal in dinosaur biogeography (англ.) // Cretaceous Research. — 2020. — ISSN 0195-6671. — doi:10.1016/j.cretres.2020.104678.
- ↑ Ostrom, J.H. 1961. Cranial morphology of the hadrosaurian dinosaurs of North America. Bulletin of the American Museum of Natural History 122: 33-186.
- ↑ Nabavizadeh, A. 2014. Hadrosauroid jaw mechanics and the functional significance of the predentary bone. In: D.A. Eberth, D.C. Evans, and P.E. Ralrick (eds.), Hadrosaurs, 467—483. Indiana University Press, Bloomington.
- ↑ Nabavizadeh, A. and Weishampel, D.B. 2016. The predentary bone and its significance in the evolution of feeding mechanisms in ornithischian dinosaurs. The Anatomical Record 299: 1358—1388.
- ↑ Prieto-Márquez, A. 2010b. Global phylogeny of hadrosauridae (Dinosauria: Ornithopoda) using parsimony and bayesian methods. Zoological Journal of the Linnean Society 159: 435—502.
- ↑ Prieto-Marquez, A., Erickson, G.M., Ebersole, J.A. 2016. A primitive hadrosaurid from southeastern North America and the origin and early evolution of 'duck-billed' dinosaurs. Journal of Vertebrate Paleontology 36 (2): e1054495.
- ↑ David B. Weishampel, Coralia-Maria Jianu. The Transsylvanian Ornithopods: Telmatosaurus transsylvanicus // Transylvanian Dinosaurs. — Baltimore, MD: the Johns Hopkins University Press, 2011. — P. 49—53.
- ↑ Nicholas R. Longrich, Xabier Pereda Suberbiola, R. Alexander Pyron, Nour-Eddine Jalil. The first duckbill dinosaur (Hadrosauridae: Lambeosaurinae) from Africa and the role of oceanic dispersal in dinosaur biogeography // Cretaceous Research. — 2020-11. — С. 104678. — ISSN 0195-6671. — doi:10.1016/j.cretres.2020.104678.
- ↑ Velasco, A. A. R. 2021. Los hadrosaurios (ornithopoda, hadrosauroidea) mexicanos: una revisión crítica. Revista Latinoamericana de Herpetología Vol.04 No.01
- ↑ Ramírez-Velasco AA (2022). "Phylogenetic and biogeography analysis of Mexican hadrosauroids". Cretaceous Research: Article 105267.
- ↑ Найдена самая старая мумия динозавра. Lenta.RU. Дата обращения: 16 сентября 2022. Архивировано 16 сентября 2022 года.
- ↑ 1 2 3 4 Rozadilla, Sebastián; Brissón-Egli, Federico; Lisandro Agnolín, Federico; Aranciaga-Rolando, Alexis Mauro; Novas, Fernando Emilio 2022. A new hadrosaurid (Dinosauria: Ornithischia) from the Late Cretaceous of northern Patagonia and the radiation of South American hadrosaurids. Journal of Systematic Palaeontology. 19 (17): 1207–1235.
- ↑ Bonaparte, J. F., Franchi, M. R., Powell, J. E. &Sepulveda, E. G. 1984. La Formacion Los Alamitos(Campaniano–Maastrichtiano) del sudeste de Rıo Negro, con descripcion de Kritosaurus australis n. sp.(Hadrosauridae). Significado paleogeografico de losvertebrados. Revista de la Asociacion Geologica Argentina, 39 (3&4), 284–299.
- ↑ Coria, R. A. 2014. South American hadrosaurs: considerations on their diversity. Pp 332–339 in D. A. Eberth & D. C. Evans (eds) Hadrosaurs. Indiana University Press, Bloomington and Indianapolis.
- ↑ 1 2 Horner J.R., Weishampel D.B., Forster C.A. Hadrosauridae. In: Weishampel DB, Dodson P, Osmólska H, editors. The Dinosauria. 2nd ed. Berkeley: University of California Press; 2004. p. 438–63.
- ↑ Nopcsa, F., 1928, «The genera of reptiles», Palaeobiologica 1:163-188
- ↑ A. Prieto-Márquez and J. R. Wagner, 2010, «Pararhabdodon isonensis and Tsintaosaurus spinorhinus: a new clade of lambeosaurine hadrosaurids from Eurasia», Cretaceous Research 30: 1238—1246
- ↑ Gates T. A., Scheetz R. A new saurolophine hadrosaurid (Dinosauria: Ornithopoda) from the Campanian of Utah. North America J Syst Palaeontol. 2015;13:711–25.
- ↑ McDonald A.T., Wolfe D.G., Freedman Fowler E.A., Gates T.A. A new brachylophosaurin (Dinosauria: Hadrosauridae) from the Upper Cretaceous Menefee Formation of New Mexico. PeerJ. 2021;9: e11084.
- ↑ Xing H., Mallon J.C., Currie M.L. Supplementary cranial description of the types of Edmontosaurus regalis (Ornithischia: Hadrosauridae), with comments on the phylogenetics and biogeography of Hadrosaurinae. PLoS ONE. 2017;12: e0175253.
- ↑ Kobayashi Y., Nishimura T., Takasaki R., Chiba K., Fiorillo A.R., Tanaka K., et al. A new hadrosaurine (Dinosauria: Hadrosauridae) from the marine deposits of the Late Cretaceous Hakobuchi Formation, Yezo Group. Japan Sci Rep. 2019;9:12389.
- ↑ Madzia D., Arbour V.M., Boyd C.A., Farke A.A., Cruzado-Caballero P., Evans D.C. The phylogenetic nomenclature of ornithischian dinosaurs. PeerJ. 2021;9: e12362.
- ↑ PBDB. paleobiodb.org. Дата обращения: 4 января 2021. Архивировано 18 сентября 2020 года.
Литература
править- Курочкин Е. Гадрозавры на Амуре // «Вокруг света», сентябрь 2006, № 9 (2792), рубрика «Спираль времени».
Ссылки
править- Утконосые динозавры // Paleoworld.ucoz.ru
- Гадрозавриды (Большие ящеры) // Dinopedia.ru
- Hadrosaurus (Гадрозавр) /вебархив/ Dinosaur.ru
- В начале палеогена ещё существовали динозавры? // Палеонтологический портал «Аммонит.ру»
- Найдены динозавры, переплывшие океан. У гадрозаврид был единственный способ попасть в Африку: пересечь океан // Вести — Наука, 6 ноября 2020